
Brian Howell博士
语言
研究项目和附属机构
研究兴趣
教育
研究抽象
大脑发育是一种精细调控的现象, 分化波由此产生不同种类的神经元, 少突胶质细胞,最后, 星形胶质细胞. 最终这些细胞, 尤其是神经元, 组织成功能网络,接收, 整合和传递信息. 我的实验室研究的是分子信号如何调节神经元从起始点向目的地的迁移, 形成突触并整合到网络中. 我们的重点之一是研究Reelin信号通路如何影响不同神经元类别在大脑中进入刻板层的定位. Reelin, 分泌的配体, 通过聚集神经元受体ApoER2和VLDLR并激活src家族激酶,诱导细胞内对接蛋白Dab1的酪氨酸磷酸化. Dab1的酪氨酸磷酸化产生包括Nckβ分子的信号复合物, Crk, CrkL和PI3K. 这些复合物调节神经元迁移时的行为, 使它们能够分层. 我们已经证明,Reelin信号影响高尔基体向神经元树突的延伸和神经元极化的能力. 我们目前正在研究这些细胞行为如何影响由Reelin信号调节的神经元定位和其他神经元特性.

图1. Reln 是出生时海马体中高尔基体伸长和定向所必需的吗. 通过Ctip2免疫染色鉴定的海马神经元通常具有延长的grasp65阳性高尔基体,延伸到根尖树突. 相反,在纯合子中 Reln 突变动物的高尔基体在细胞核近端弯曲.
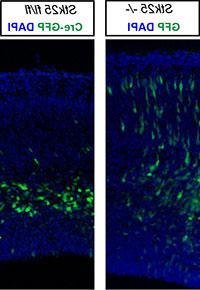
图2. Stk25是一种磷酸化Tau表型的基因修饰因子 Dab1 突变小鼠,影响神经元迁移. 与本构相反 Stk25 基因敲除,一个条件等位基因的急性失活 Stk25 绿色荧光蛋白-Cre在E14的表达.5引起E17明显的异常神经元迁移.5.
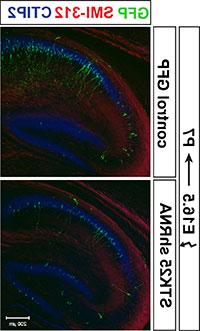
图3. 击倒的 Stk25 导致树突发生异常. Stk25 shRNA在E16海马中的表达.5导致顶端和基部树突之间的树突不对称减少.
We have recently exp和ed our focus to include genetic causes of autism spectrum disorder (ASD); specifically RELN 其中40多种突变已在ASD患者中被发现. 我们发现突变富集在Reelin亚重复结构域核心的Arg-X-Arg基序中. 配对精氨酸残基的函数是未知的. 因此, 我们正在努力解决Reelin生物合成和信号传导中该基序突变的后果,最终目标是确定它们如何导致ASD.
选定的出版物
Sutt年代, Cansby E, 保罗一, Amrutkar米, 努涅斯·杜兰E, Kulkarni纳米, 斯塔尔米, 博伦J, Laurencikiene J, 豪厄尔BW, Enerbäck S, Mahlapuu M. STK25调节脂肪组织的氧化能力和代谢效率. J性. 2018年5月24日. pii:乔- 18 - 0182. doi: 10.1530 /乔- 18 - 0182. [Epub预印]PMID:29794231
拉默特,维.B.米德尔顿,F.A.潘,J.奥尔森,E.C. 和 豪厄尔,B.W. (2017) 的 新创 自闭症谱系障碍 RELN R2290C突变减少Reelin分泌并增加蛋白二硫异构酶的表达. J Neurochem. 142:89-102. doi: 10.1111 / jnc.14045
拉默特DB, 豪厄尔BW. 自闭症谱系障碍中的RELN突变. 前细胞神经科学. 2016年3月31日;10:84. doi: 10.3389 / fncel.2016.00084. eCollection 2016. 审查.PMID: 27064498
Abadesco AD, Cilluffo M, Yvone GM, Carpenter EM, 豪厄尔BW菲尔普斯体育. 在成人脑和脊髓中发现新的表达disabled -1的神经元. 【推荐最近最火的赌博软件】. 2014年2月,39 (4):579 - 92. doi: 10.1111 / ejn.12416. 2013年11月19日.PMID: 24251407
Matsuki T, Chen J, 豪厄尔BW. 丝氨酸-苏氨酸激酶Stk25的急性失活会破坏神经元迁移. 神经系统开发. 2013年11月13日;8:21. doi: 10.1186/1749-8104-8-21.PMID: 24225308
特谢拉厘米, 克隆亚麻毫米, Masachs N, 张H, Lagace直流, 马丁内斯一, Reillo我, 段X, 博世C, Pujadas L, Brunso L, 歌H, Eisch AJ, 博雷利V, 豪厄尔BW,家长JM,索里亚诺E. 细胞自主失活的reelin通路损害海马成体神经发生. J >. 2012年8月29日;32(35):12051-65.PMID: 22933789
Matsuki T, Zaka M, Guerreiro R, van der Brug MP, Cooper JA, Cookson MR, Hardy JA, 豪厄尔BW. Stk25作为dab1突变小鼠Tau磷酸化基因修饰因子的鉴定. 《推荐最近最火的赌博软件》. 2012;7(2):e31152. doi: 10.1371 /杂志.玉米饼.0031152. 2012年2月15日.PMID: 22355340
Matsuki T, Matthews RT, Cooper JA, van der Brug MP, Cookson MR, Hardy JA, Olson EC, 豪厄尔BW. Reelin和stk25在神经元极化和树突高尔基体部署中具有相反的作用. 细胞. 2010年11月24日;143(5):826-36. doi: 10.1016/j.细胞.2010.10.029.PMID: 21111240
Matsuki T, Pramatarova A, 豪厄尔BW. 降低Crk和CrkL的表达可阻断reelin诱导的树突发生. [J]细胞科学. 2008年6月1日;121(11):1869-75. doi: 10.1242 / jcs.027334. 2008年5月13日.PMID: 18477607
Pramatarova A, Chen K, 豪厄尔BW. APP和Dab1基因之间的遗传相互作用影响大脑发育. Mol细胞神经科学. 2008年1月,37 (1):178 - 86. Epub 2007 9月26日.PMID: 18029196
Pramatarova A, Ochalski PG, Lee CH, 豪厄尔BW. 小鼠disabled 1调节果蝇眼模型中神经元的核位置. Mol细胞生物学. 2006年2月,26 (4):1510 - 7.PMID: 16449660
豪厄尔,B.W.赫里克,T.M.希尔德布兰德,J.D.张,Y.库珀,J.A. (2000). Dab1酪氨酸磷酸化位点在小鼠大脑发育过程中传递位置信号. 现代生物学10:877-885. PMID: 10959835
豪厄尔,B.W.赫里克,T.库珀,J.A. (1999) reelin诱导的神经元定位过程中Disabled 1的酪氨酸磷酸化. 基因开发.13 (6):643-648
豪厄尔,B.W.霍克斯,R.索里亚诺,P.库珀,J.A. (1997). 发育中的大脑神经元位置受小鼠残疾调节. 自然科学学报,39 (3):733-737.